Популярные статьи

Популярное
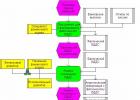
Особенности ведения бухгалтерии курьерских служб Отражены расходы на доставку товаров покупателям силами сторонней курьерской службы
Вести бухгалтерский учёт обязаны все юридические лица вне зависимости от размера бизнеса и организационно-правовой формы. Зачастую...